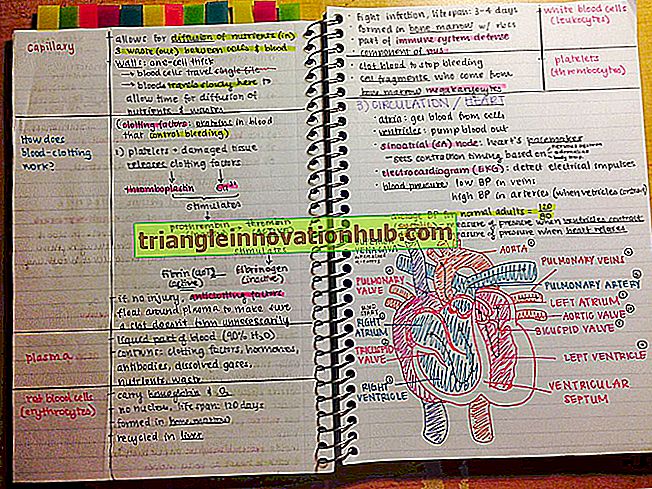
3 Tipos de Reprodução Sexual que ocorre em Bactérias (1869 Palavras)
Tipos de reprodução sexual que ocorre nas bactérias são os seguintes:
Observações citológicas e estudos genéticos indicam algo como a reprodução sexual, envolvendo a fusão de duas células diferentes e uma transferência de fatores hereditários ocorre em bactérias, embora com pouca freqüência. A recombinação genética ocorre nas bactérias que foram cuidadosamente estudadas e presumivelmente também ocorre em outras espécies.

Imagem Cortesia: upload.wikimedia.org/wikipedia/commons/thumb/c/c7/Caduco.jpg/1280px-Caduco.jpg
Uma das espécies de bactérias mais intensamente estudadas, a Escherichia coli, demonstrou ter sexo - algumas atuam como machos e transferem informações genéticas pelo contato direto com as fêmeas. Essa capacidade de transferir genes é regulada por um fator de fertilidade F + que pode ser transferido para uma fêmea, convertendo-a em um macho.
As células bacterianas vegetativas usuais são haplóides e, na reprodução sexual, parte ou todo o cromossomo passa da célula masculina para a célula feminina, produzindo uma célula, isto é, parcialmente ou completamente diploide. O cruzamento ocorre então entre o cromossomo feminino e o cromossomo ou fragmento masculino, seguido por um processo de segregação que produz células progênicas haplóides.


1. transformação bacteriana:
A transferência genética em bactérias também ocorre por transformação, na qual a molécula de DNA da célula doadora, quando liberada por sua desintegração, é absorvida por outra célula receptora e seus descendentes herdam alguns caracteres da célula doadora. Quando diferentes cepas de bactérias são encontradas em um estado misto, seja em cultura ou na natureza, alguns dos descendentes resultantes possuem uma combinação de caracteres das cepas parentais. Esse fenômeno é conhecido como recombinação.
O fenômeno da transformação foi registrado pela primeira vez por Griffith (1928). Avery, Macleod e McCarty (1944) demonstraram que o princípio transformador é o DNA na seqüência de eventos na transformação bacteriana.
As linhas de investigação que levaram a uma compreensão da natureza química do material genético surgiram de um estudo do organismo pestilento Diplococcus pneumoniae. Esta bactéria causa pneumonia nos machos. Em 1928, Frederick Griffith descobriu que existem duas cepas de D. pneumoniae, uma que forma colônias suaves protegidas por uma cápsula, e outra que formava colônias irregulares ou ásperas sem cápsula quando cultivadas em meio adequado em placas de petri.
Quando injetada em camundongos (A), apenas células lisas capsuladas (virulentas) produziram a doença, mas não as células rugosas não virulentas (B). Por outro lado, quando as células lisas encapsuladas (virulentas) mortas pelo calor foram misturadas com células ásperas não virulentas (D) e depois foram injectadas nos ratinhos, a doença foi produzida. Isso mostra que alguns fatores das células lisas encapsuladas mortas, converteram as células ásperas não virulentas vivas em células vivas encapsuladas (virulentas), (veja fig. 2.16).
Em 1944, Avery, McCarty e Macleod apoiaram o experimento de Griffith pela explicação molecular. Eles descobriram que o DNA isolado do calor matou as células lisas, quando adicionado às células ásperas, mudou seu caráter superficial de áspero para suave, e também as tornou virulentas.
Por este experimento, foi demonstrado que o DNA era o material genético responsável por induzir o caráter suave das células e sua propriedade de virulência em camundongos. Sua experiência provou que a transformação bacteriana envolve a transferência de uma parte do DNA da bactéria morta (ou seja, doador) para a bactéria viva (isto é, receptor), que expressa o caráter de célula morta e, portanto, é conhecida como recombinante.

O agente infectante viral é o DNA:
Um bacteriófago (vírus T 2 ) infecta a bactéria Escherichia coli. Após a infecção, o vírus se multiplica e os fagos T 2 são liberados com a lise das células bacterianas. Como sabemos, o fago T 2 contém DNA e proteínas. Agora surge a pergunta, qual dos dois componentes tem a informação para programar a multiplicação de mais partículas virais.
Para resolver este problema, Hershey e Chase (1952) conceberam uma experiência com duas preparações diferentes de fago T2. Em uma preparação eles fizeram a proteína parte radioativa e na outra preparação o DNA foi feito radioativo. Depois disso, uma cultura de E. coli foi infectada por estas duas preparações fágicas. Imediatamente ap� a infec�o e antes da lise das bact�ias, as c�ulas de E. coli foram agitadas suavemente num misturador de modo a que as part�ulas f�icas aderentes fossem soltas e depois a cultura fosse centrifugada. Com o resultado, os pellets mais pesados de células bacterianas infectadas foram depositados no fundo do tubo. As partículas virais mais leves e as partículas que não entraram nas células bacterianas foram encontradas no sobrenadante. Verificou-se que quando o fago T2 com ADN radioactivo foi utilizado para infectar E. coli na experiência, o sedimento bacteriano mais pesado era também radioactivo. Por outro lado, quando foi utilizado o fago T 2 com protea radioactiva, o sedimento bacteriano tinha muito pouca radioactividade e a maior parte da radioactividade foi encontrada no sobrenadante. este


que é o DNA viral e não a proteína que contém informações para a produção de mais partículas de fago T 2, portanto o DNA é material genético. No entanto, em alguns vírus (por exemplo, vírus da influenza, vírus da influenza e vírus da poliomielite), o RNA serve como material genético (ver figura 2.17).

Hershey e Chase conduziram dois experimentos. Numa expericia, E. coli foi administrada num meio contendo o radioisopo S35 e na outra expericia, E. coli foi cultivada num meio contendo o radio-isopo P 32 . Nestas expericias, as culas de E. coli foram infectadas com fago T2 libertado de culas de E. coli cultivadas em meio S35 com S35 na sua protea capsde e as do meio P32 tinham P 32 no seu ADN.
Quando estes fagos foram utilizados para infectar novas culas de E. coli em meio normal, as culas bacterianas que tinham infeco por fagos marcados com S35 mostraram a radioactividade na sua parede celular e n no citoplasma. Considerando que as bactérias infectadas com fagos marcados com P32 mostraram a condição inversa.
Assim, pode-se dizer que, quando o fago T 2 infecta a célula bacteriana, sua cápside protéica permanece fora da célula bacteriana, mas seu DNA entra no citoplasma da bactéria. Quando as células infectadas das bactérias são lisadas, novas partículas virais completas (fagos T 2 ) são formadas. Isso prova que o DNA viral carrega as informações para a síntese de mais cópias de DNA e capsídeos de proteína. Isso mostra que o DNA é material genético (veja a figura 2.19).
2. Transdução bacteriana:
A transferência genética em bactérias é conseguida por um processo conhecido como transdução. O experimento de Lederberg e Zinder (1952) no tubo U Salmonella typhimurium indicou que os vírus ou fagos bacterianos são responsáveis pela transferência de material genético de um para outro lisogênico e

fagos líticos. Assim, o hospedeiro adquire um novo genótipo. A transdução foi demonstrada em muitas bactérias.
Neste processo, a molécula de DNA que transporta os caracteres hereditários da bactéria doadora está sendo transferida para a célula receptora através da ação da partícula do fago. Neste processo, poucos caracteres intimamente ligados podem ser transferidos por cada partícula. Assim, o bacteriófago provoca mudanças genéticas nas bactérias que sobrevivem ao ataque do fago.

Quando uma célula bacteriana está sendo infectada com um vírus temperado, começa o ciclo lítico ou a lisogenia. Posteriormente, o DNA do hospedeiro se divide em pequenos fragmentos juntamente com a multiplicação do vírus. Alguns desses fragmentos de DNA são incorporados com as partículas do vírus que se transformam em uma. Quando as bactérias lisam essas partículas junto com partículas normais de vírus são liberadas

quando se permite que esta mistura de partículas de transdução e de vírus normais infecte a população de células receptoras, a maioria das bactérias é infectada com partículas virais normais e com o resultado ocorre novamente lisogenia ou ciclo lítico. Algumas bactérias são infectadas com partículas de transdução, a transdução ocorre e o DNA das partículas do vírus sofre recombinações genéticas com o DNA bacteriano. (Veja as figs. 2.20 e 2.21).
3. conjugação bacteriana:
Wollman e Jacob (1956) descreveram a conjugação em que duas bactérias ficam lado a lado por até meia hora. Durante este perdo de tempo, uma poro de material genico passa lentamente de uma bactia designada como macho para um receptor designado como fea. Isto foi estabelecido que o material masculino entrou na fêmea em uma série linear.
A recombinação genética entre células doadoras e receptoras ocorre da seguinte maneira: O DNA de Hfr, depois de deixar uma parte no fragmento para a célula receptora, novamente é reformulado de maneira circular. Na linhagem F, a recombinação genética ocorre entre o fragmento doador e o DNA receptor. A transferência de genes é um processo sequencial e uma determinada cepa de Hfr sempre doa genes em uma ordem específica. Um DNA de dador de cadeia simples (factor F) está integrado no cromossoma hospedeiro com a ajuda da enzima nuclease (ver Figs. 2.21 e 2.22).
Na conjugação bacteriana, a transferência de material genético (DNA) ocorre por contato célula a célula de células doadoras e receptoras. Durante o processo de conjugação grande porção do genoma é transferida, enquanto na transformação e transdução apenas pequeno fragmento de DNA é transferido. O processo de conjugação foi descoberto por Lederberg e Tatum (1944) em uma única cepa de Escherichia coli. Conjugação também foi demonstrada em Salmonella, Pseudomonas e Vibrio.

Em conjugação, a transferência unidirecional de material genético ocorre da doação à cepa receptora. As cepas doadoras e receptoras são sempre determinadas geneticamente. A estirpe receptora é designada como F, enquanto que as estirpes dadoras são de dois tipos e são designadas como F + e H fr (alta frequência de recombinação). Se a cepa doa apenas uma pequena porção de seu genoma, ela é chamada F + e, se doar grande quantidade de genoma, é chamada de H fr. Esses fatores F + e H fr são chamados de episomes.
As cepas F + e Hfr são caracterizadas pela presença de estruturas semelhantes a flagelos específicos, o chamado sexo pilus. O pilus sexual está ausente nas cepas F + e é responsável pelo acasalamento bacteriano. Pilos de sexo de F + e H fr tocam o tipo oposto de células de acasalamento especificamente para transferir o material genético.
O pilus sexual tem um orifício de 2, 5µm de diâmetro que é grande o suficiente para uma molécula de DNA passar através dele no sentido do comprimento. No momento do emparelhamento do DNA da estirpe de H fr (dador) é transferido para a estirpe F (recipiente) imediatamente. O DNA circular das células H fr abre e se replica, mas durante a transferência, uma fita de DNA é sintetizada de novo, enquanto a outra é derivada de uma fita pré-existente de H fr. Após a transferência do DNA, ambas as células são separadas umas das outras.
O DNA de H fr, depois de separar o seu fragmento para a célula receptora, volta a ser reformado de maneira circular. Na linhagem F, a recombinação genética ocorre entre o fragmento doador eo DNA receptor. A transferência gênica é um processo seqüencial em que uma determinada cepa H fr sempre doa genes em uma ordem específica. Se as estirpes F - e H fr forem misturadas numa suspensão, diferentes genes numa sequência de tempo são transferidos do genoma de H fr para F - estirpe. Os genes que entram precocemente aparecem sempre em maior porcentagem das recombinações do que os genes que entram mais tarde (ver figs. 2.22, 2.23 e 2.24).

A conjugação resulta em vários recombinantes em uma suspensão de células F + e H fr. Esses recombinantes são variáveis em sua constituição genotípica e também em sua expressão fenotípica. Esses recombinantes são inteiramente novos e diferentes de seus pais.